细胞分化
脊椎动物和人体由约200多种不同类型的细胞组成,这些由单个受精卵产生的细胞,在形态结构、生化组成和功能等方面均有明显的差异,形成这种稳定性差异的过程称为细胞分化(cell differentiation)。
一、多细胞生物个体发育过程与细胞分化潜能
细胞分化是个体发育过程中细胞在结构和功能上发生差异的过程。多细胞生物的个体发育一般包括胚胎发育和胚后发育两个阶段,细胞分化贯穿于个体发育的全过程,其中胚胎期最明显。
- 胚胎发育:包括卵裂、囊胚、原肠胚、神经轴胚期及器官发生等阶段,衍生出与亲代相似的幼小个体;
- 胚后发育:幼体从卵膜孵化出或从母体分娩以后,经幼年、成年、老年直至衰老、死亡的过程。
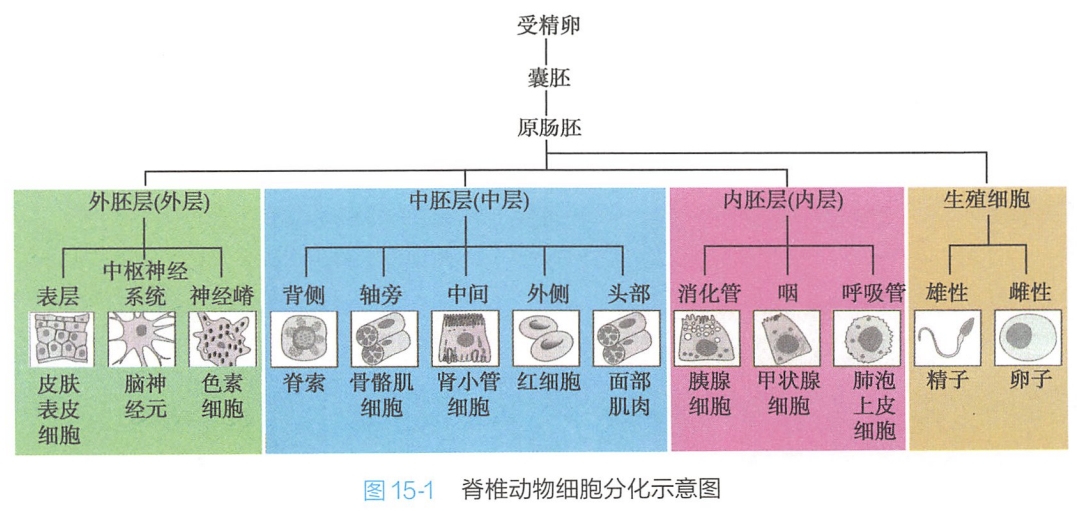
(一)动物和人类胚胎的三胚层代表不同类型细胞的分化去向
卵细胞在受精后立刻进入反复的有丝分裂阶段,这一快速的分裂时期称为卵裂(cleavage)。
动物早期胚胎发育中受精卵经过卵裂被分割成许多小细胞,这些小细胞组成的中空球形体被称为囊胚(blastula)。
囊胚形成后,便进入原肠形成(gastrulation)期。原肠胚期之前,细胞间并无可识别的明显差异。在原肠胚期,产生了内、中、外三个胚层,它们具有不同的发育和分化去向:
- 内胚层(endoderm)将发育为消化道及其附属器官、睡液腺、胰腺、肝脏以及肺等的上皮成分;
- 中胚层(mesoderm)将发育成骨骼、肌肉、纤维组织和真皮,以及心血管系统和泌尿系统;
- 外胚层(ectoderm)则形成神经系统、表皮及其附属物(图15-1)。
(二)细胞分化的潜能随个体发育进程逐渐“缩窄”
随着个体发育进程,来源于单一受精卵的细胞逐渐产生出形态结构、功能和生化特征各不相同的细胞类群,其结果在空间上细胞产生差异,在时间上同一细胞和从前的状态有所不同。
研究表明,两栖类动物在囊胚形成之前的卵裂球细胞、哺乳动物桑葚胚的8细胞期之前的细胞和其受精卵一样,均能在一定条件下分化发育成为完整的个体。通常将具有这种特性的细胞称为全能性细胞(totipotent cell)。
在三胚层形成后,由于细胞所处的空间位置和微环境的差异,细胞的分化潜能受到限制,各胚层细胞只能向本胚层组织和器官的方向分化发育,而成为多能细胞(pluripotent cell)。
经过器官发生(organogenesis),各种组织细胞的命运最终确定,呈单能(unipotency)化。这种在胚胎发育过程中,细胞逐渐由“全能”到“多能”,最后向“单能”的趋向,是细胞分化的一般规律。
近些年从囊胚内细胞团(inner cell mass,ICM)中分离到的胚胎干细胞(embryonic stem cell,ES细胞),它们具有分化成熟为个体中所有细胞类型的能力,但不能分化为胎盘和其他一些发育时所需的胚外组织,这种早期胚胎细胞被称为多能干细胞(pluripotent stem cell,multipotential stem cell)。因此,从干细胞角度看细胞分化,细胞呈现出由“全能干细胞”到“多能干细胞”,最后向“定向干细胞”和“前体细胞”的趋向(图15-2)。
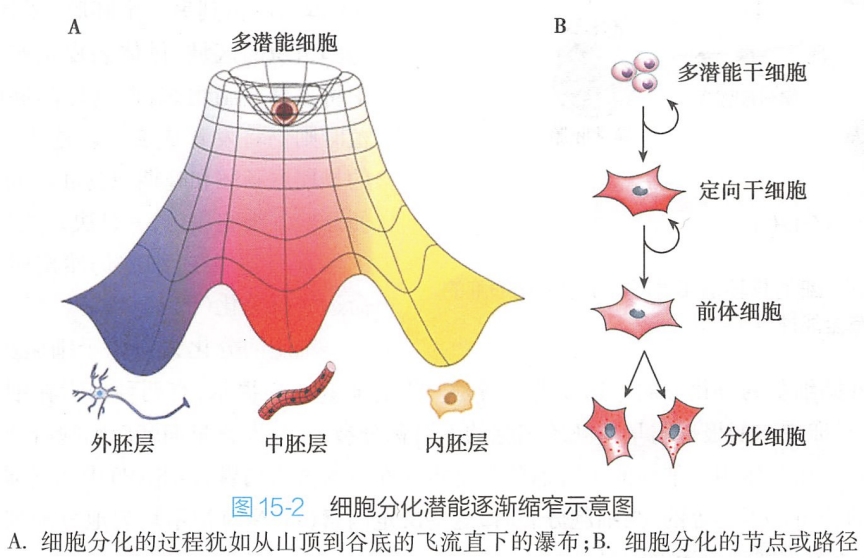
应当指出的是,大多数植物和少数低等动物(如水螅)的体细胞仍具有全能性;而在高等动物和人类,至成体期,除一些组织器官保留了部分微分化的组织干细胞之外,其余均为终末分化细胞。
(三)终末分化细胞的细胞核具有全能性
动物受精卵子代细胞的全能性随其发育过程逐渐受到限制而变窄,即由全能性细胞转化为多能和单能干细胞,直至分化为终末细胞。但在细胞核则完全不同,终末分化细胞的细胞核仍然具有全能性,谓之全能性细胞核(totipotent nucleus)。
1997年,将成年绵羊乳腺上皮细胞的细胞核移植到另一只羊的去核卵细胞中,成功地克隆出世界上第一只哺乳动物一“多莉”羊。实验结果表明,已特化的体细胞的细胞核仍保留形成正常个体的全套基因,具有发育成一个有机体的潜能。
二、细胞决定与细胞分化
(一)细胞决定先于细胞分化并制约着分化的方向
在个体发育过程中,细胞在发生可识别的分化特征之前就已经确定了未来的发育命运,只能向特定方向分化的状态,称之为细胞决定(cell determination)。
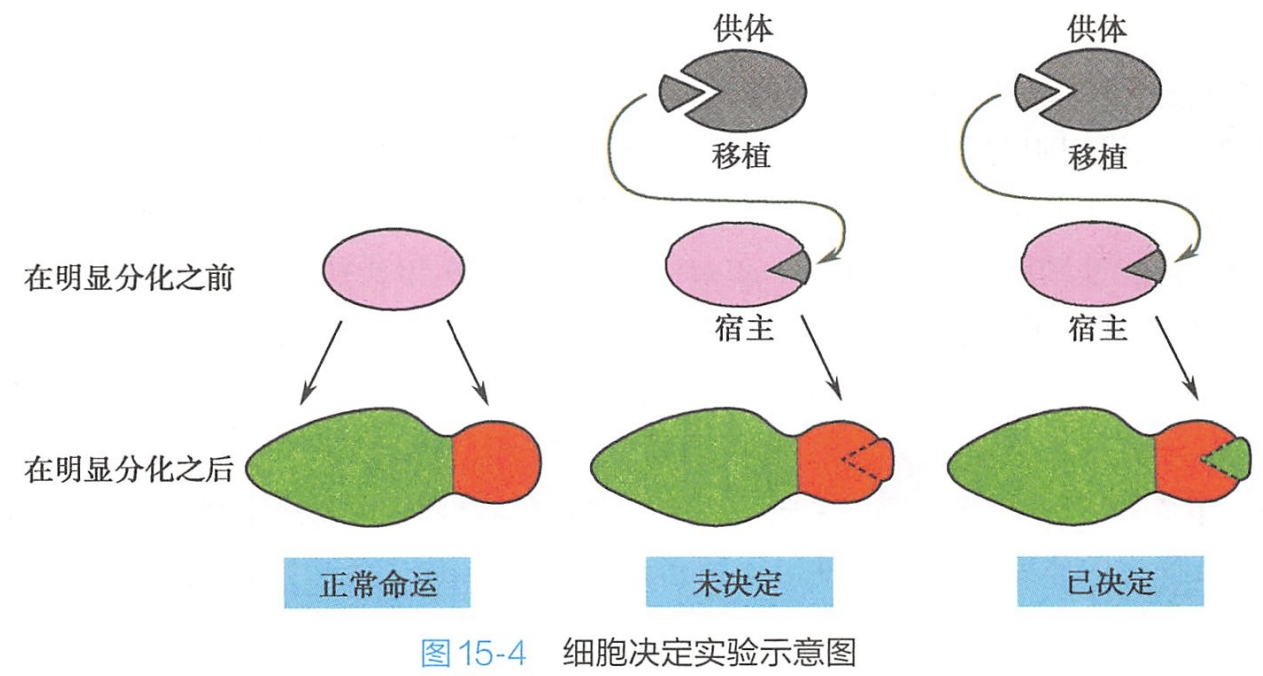
在原肠期的内、中、外三胚层形成时,虽然在形态上看不出有什么差异,但此时形成各器官的预定区已经确定,每个预定区决定了它只能按一定的规律发育分化成特定的组织、器官和系统。细胞决定可通过胚胎移植实验(grafting experiment)予以证明。
例如在两栖类胚胎,如果将原肠胚早期预定发育为表皮的细胞(供体),移植到另一个胚胎(受体)预定发育为脑组织的区域,供体表皮细胞在受体胚胎中将发育成脑组织,而到原肠胚晚期阶段移植时则仍将发育成表皮。这表明,在两栖类的早期原肠胚和晚期原肠胚之间的某个时期便开始了细胞决定,一旦决定之后,即使外界的因素不复存在,细胞仍然按照已经决定的命运进行分化(图15-4)。
细胞的分化去向源于细胞决定,是什么因素决定了胚胎细胞的分化方向?迄今尚不清楚。现有研究资料提示,有两种因素在细胞决定中起重要作用:
- 一是卵细胞的极性与早期胚胎细胞的不对称分裂,细胞的不对称分裂是指存在于核酸蛋白颗粒(RNP)中的转录因子RNA在细胞质中的分布是不均等的,当细胞分裂时,这些决定因素(RNA)被不均匀地分配到两个子细胞中,结果造成两个子细胞命运的差异。
- 二是发育早期胚胎细胞处的位置差异及胚胎细胞间的相互作用。
(二)细胞决定具有遗传稳定性
细胞决定表现出遗传稳定性,典型的例子是果蝇成虫盘细胞的移植实验。
成虫盘是幼虫体内已决定的尚未分化的细胞团,在幼虫发有的变态期之后,不同的成虫盘可以逐渐发育为果蝇的腿、翅、触角等成体结构。如果将成虫盘的部分细胞移植到一个成体果蝇腹腔内,成虫盘可以不断增殖并一直保持于未分化状态,即使在果蝇腹腔中移植多次、经历1800代之后再移植到幼虫体内,被移植的成虫盘细胞在幼虫变态时,仍能发育成相应的成体结构。这说明果蝇成虫盘细胞的决定状态是非常稳定并可遗传的。人们在认识到细胞决定的稳定性和可遗传性的同时,也开始探索细胞决定的可逆性。在果蝇研究中发现,有时某种培养的成虫盘细胞会出现不按已决定的分化类型发育,而是生长出不是相应的成体结构,发生了转决定(transdetermination)。
探讨转决定的发生机制对了解胚胎细胞命运的决定具有重要意义。近年来有关细胞命运决定的主要研究策略:一是利用模式生物,分析选择性干预(如基因敲除)早期胚胎中某个基因的表达对内、中、外三胚层形成的影响;二是基于ES细胞,寻找决定ES细胞向三胚层细胞分化的决定因子。
三、细胞分化的可塑性
一般地,细胞分化具有高度的稳定性。细胞分化的稳定性(stability)是指在正常生理条件下,已经分化为某种特异的、稳定类型的细胞一般不可能逆转到未分化状态或者成为其他类型的分化细胞。
细胞分化的可塑性(plasticity)是指已分化的细胞在特殊条件下重新进入未分化状态或转分化为另一种类型细胞的现象。
(一)去分化
一般情况下,细胞分化过程是不可逆的。然而在某些条件下,分化了的细胞也不稳定,其基因活动模式也可发生可逆性的变化,而又回到未分化状态,这一变化过程称为去分化(dedifferentiation)。
在动物和人类,体细胞部分去分化的例子较多(如蝾螈肢体再生时形成的胚芽细胞及人类的各种肿瘤细胞等),但体细胞通常难以完全去分化而成为全能性细胞。
近些年干细胞领域研究的进步,已能通过人为手段成功地将分化成熟细胞逆转为未分化细胞,此即细胞重编程(cellular reprogramming)。前面讲到的基于体细胞核移植技术进行的动物克隆实验就是细胞重编程的例子。然而细胞重编程概念的真正形成和发展,源于2006年日本科学家S.Yamanaka(山中伸弥)等人的工作。山中伸弥借助逆转录病毒载体,将四个转录因子(Oct3/4、Sox2、c-Myc、Klf4)基因导入小鼠皮肤成纤维细胞(fibroblast)中,可以使来自胚胎小鼠或成年小鼠的成纤维细胞获得类似胚胎干细胞(embryonic stem cell)的多能性。一般将通过这种方法获得的多能性细胞称为诱导多能干细胞(induced pluripotent stem cells,iPS细胞)
(二)转分化
在高度分化的动物细胞中还可见到另一种现象,即从一种分化状态转变为另一种分化状态,这种情况称为转分化(transdifferentiation)。
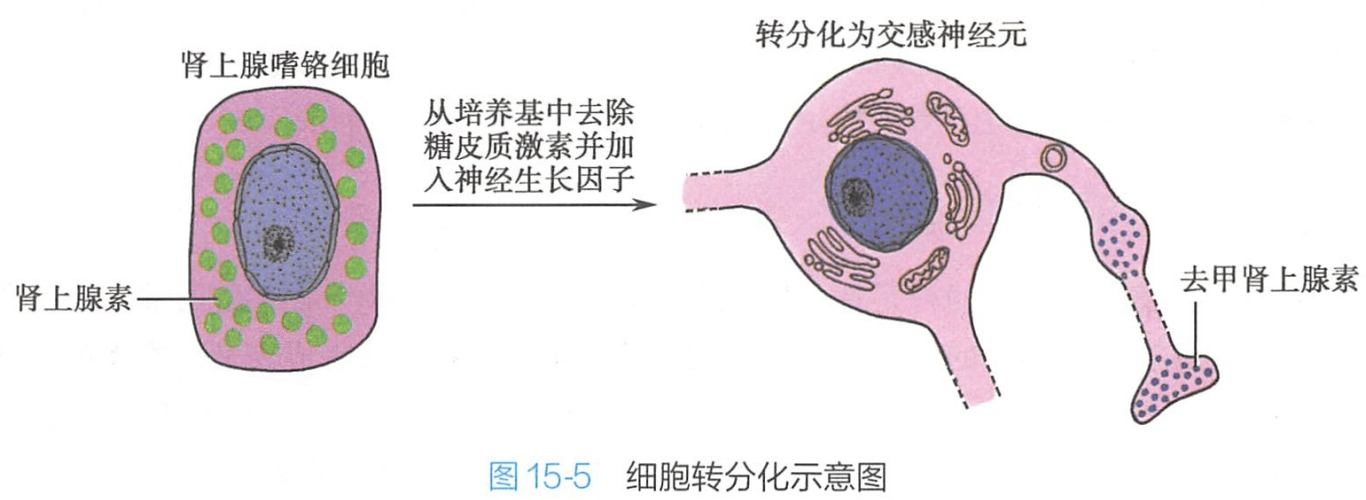
细胞通过转分化能形成一种发育相关的细胞类型。典型的例子可见于肾上腺的嗜铬细胞。体积较小的嗜铬细胞源于神经嵴并且分泌肾上腺素入血。在培养条件下,加入糖皮质激素可以维持嗜铬细胞的表型,但是当去除甾体激素并在培养基中加入神经生长因子之后,嗜铬细胞转分化成交感神经元,这些神经元比嗜铬细胞大,带有树突样和轴突样突起,并且分泌去甲肾上腺素而非肾上腺素(图15-5)。
另一个转分化的例子是,如果把鸡胚视网膜色素上皮细胞置于特定培养条件下,细胞色素则渐渐消失并且细胞开始呈现晶体细胞的结构特征,并产生晶体特异性蛋白一晶体蛋白。
上面的两个例子,通过转分化生成了一种发育相关的细胞类型:交感神经元和嗜铬细胞均来源于神经嵴;色素细胞和晶体细胞均来源于外胚层并且涉及眼的发育。通过转分化形成不同发育类型细胞的例子也较常见。
例如,水母横纹肌可由一种细胞类型连续转分化成两种不同类型的细胞。离体的横纹肌与其相关的细胞外基质共同培养时,可以保持横纹肌的状态。在用能降解细胞外基质的酶处理培养组织之后,细胞将形成一个聚合体,其中有些细胞在 1~2天内转分化为平滑肌细胞,继续培养时,可形成第二种类型的细胞——神经元。
必须指出的是,无论是动物还是植物,细胞分化的稳定性是普遍存在的,可以认为分化具有单向性、序列性和终末性(一般情况下都会到达分化的目标终点,成为终末分化细胞),而去分化是逆向运动,转分化是转序列运动。发生细胞的转分化或去分化是有条件的:
- 细胞核必须处于有利于分化逆转环境中,
- 分化能力的逆转必须具有相应的遗传物质基础。通常情况下,细胞分化的逆转易发生于具有增殖能力的组织中。
四、细胞分化的时空性
在个体发育过程中,多细胞生物细胞既有时间上的分化,也有空间上的分化。一个细胞在不同的发育阶段可以有不同的形态结构和功能,即时间上的分化;
同一种细胞的后代,由于每种细胞所处的空间位置不同,其环境也不一样,可以有不同的形态和功能,即空间上的分化。
在高等动植物个体胚胎发育过程中,随着细胞数目的不断增加,细胞的分化程度越来越复杂,细胞间的差异也越来越大;同一个体的细胞由于所处的空间位置不同而确定了细胞的发育命运,出现头与尾、背与腹等不同。
这些时空差异为形成功能各异的多种组织和器官提供了基础。
五、细胞分裂与细胞分化
细胞分裂和细胞分化是多细胞生物个体发育过程中的两个重要事件,两者之间有密切的联系。通常细胞在增殖(细胞分裂)的基础上进行分化,而早期胚胎细胞的不对称分裂所引起的细胞质中转录因子的差异制约着细胞的分化方向和进程。
细胞分化发生于细胞分裂的G1期,在早期胚胎发育阶段特别是卵裂过程中,细胞快速分裂,G1期很短或几乎没有G1期,此时细胞分化减慢。细胞分裂旺盛时分化变缓,分化较高时分裂速度减慢是个体生长发育的一般规律。
例如哺乳动物的表皮角化层细胞等终末细胞分化程度较高,分裂频率明显减慢,而高度分化的细胞,如神经元和心肌细胞则很少分裂或完全失去分裂能力。